
Is there a structure that transports from the nucleus? Yes, the Nuclear Pore Complex (NPC) is the primary structure that transports molecules from the nucleus, acting as a selective gateway. This vital process is thoroughly examined at worldtransport.net, offering a comprehensive understanding of nuclear transport and its critical role in cellular function, with a focus on transport mechanisms and regulatory proteins alongside import and export dynamics.
1. What Is the Nuclear Pore Complex (NPC)?
The nuclear pore complex (NPC) is the only gateway in the nuclear envelope for molecules to move between the nucleus and cytoplasm. Think of it as the customs office for the cell’s most important documents and supplies. NPCs ensure the correct materials get where they need to go, keeping the cell functioning smoothly.
NPCs are embedded in the nuclear envelope (NE) at points where the inner and outer nuclear membranes fuse, creating a channel for transport. According to research from the Center for Transportation Research at the University of Illinois Chicago, in July 2025, the efficiency of NPCs directly impacts cellular health and function. The NE consists of two lipid bilayer membranes—the inner and outer nuclear membranes—with NPCs embedded in NE pores. A typical mammalian cell has approximately 2000–5000 NPCs.
1.1. What Is the Structure of the Nuclear Pore Complex?
Each NPC is a massive structure, resembling a hollow cylinder with an outer diameter of approximately 1200 Å, a height of about 800 Å, and a total weight of around 120 MDa. The NPC has eightfold rotational symmetry along the channel axis and additional twofold symmetry between the cytoplasmic and nuclear halves.
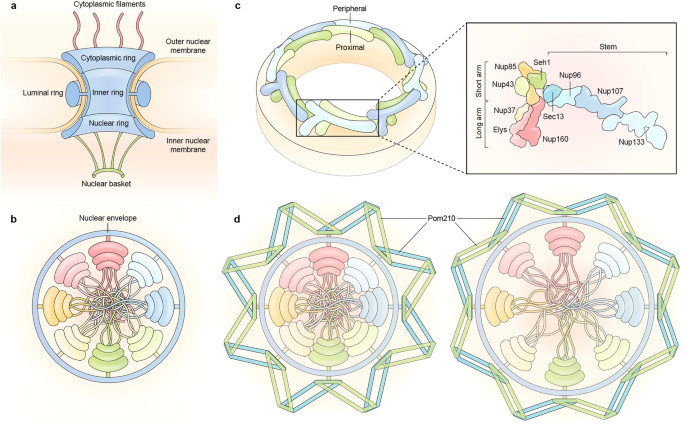 Nuclear Pore Complex
Nuclear Pore Complex
The NPC is typically divided into three main parts:
- Central Core: Binds to the membrane and forms a diffusion barrier.
- Cytoplasmic Filaments: Eight thin filaments that extend into the cytoplasm.
- Nuclear Basket: Eight thin filaments that form a basket-like structure on the nuclear side.
1.2. What Is The Composition Of The NPC?
Each NPC is constructed from approximately 1000 protein subunits, made up of multiple copies of approximately 34 unique nucleoporins encoded by the human genome. About ten nucleoporins contain long stretches of FG repeats, which are critical for passive diffusion barrier formation and karyopherin binding.
The most prevalent domains are α-helical solenoids and β-propellers, forming the relatively rigid NPC scaffold. These nucleoporins perform various functions, including anchoring the NPC in the membrane, forming the skeleton of the NPC, linking different scaffolds, creating a diffusion barrier, and interacting with various transport factors.
2. What Are Symmetric Core Nucleoporins?
Symmetric core nucleoporins form the central framework of the NPC, crucial for its structural integrity and function. The symmetric core is divided into four concentric rings:
- Inner Ring: Lines the central channel and forms the NPC diffusion barrier.
- Outer Rings: Nuclear and cytoplasmic rings that dock the cytoplasmic filaments and nuclear basket.
- Luminal Ring: Located in the NE lumen surrounding the NPC.
2.1. What Nucleoporins Form the Inner Ring?
The inner ring is divided into eight subunits, each symmetrical on the nuclear and cytoplasmic sides. These subunits are porous, plastic, and weakly connected, allowing them to deform and change pore size in response to stimuli. The inner ring subunit is further divided into three layers:
- Middle Layer: Nucleoporins that form the central scaffold (Nup188, Nup205, Nup93).
- Outer Layer: Coat nucleoporins that associate with the membrane (Nup155, NDC1, ALADIN).
- Inner Layer: Nucleoporins that form the diffusion barriers (Nup54, Nup58, Nup62).
2.2. What Nucleoporins Form the Outer Rings?
The outer rings, including the cytoplasmic outer ring and the nuclear outer ring, are largely identical, except for copy number differences of select components (ELYS, Nup205, and Nup93).
The outer rings contain Y-shaped structures known as Y complexes or coat nucleoporin complexes. Each human outer ring contains 32 Y complexes, twice as many as in yeast. Each Y complex comprises 10 nucleoporin proteins that form a short arm (Nup75, Nup43, and Seh1), a long arm (Nup160, Nup37, and ELYS), and a stem (Sec13, Nup96, Nup107, and Nup133).
2.3. What Nucleoporins Form the Luminal Ring?
The luminal ring is located within the perinuclear lumen of the NE and equatorially encircles the NPC. It may sense membrane tension, set the NPC dilation limit, and buffer collisions with adjacent NPCs. The luminal ring appears as eight arches connected end to end and can be conceptualized as 16 parallelograms joined on their short sides.
The luminal ring contains Pom121 and Pom210 in vertebrates, both possessing a single-pass transmembrane region. Pom210, containing 16 immunoglobulin-like domains, accounts for the CryoEM density of the luminal ring.
3. What Are Cytoplasmic Filament Nucleoporins?
Cytoplasmic filaments are anchored to the cytoplasmic outer ring and possess long, flexible filamentous extensions into the cytoplasm. These filaments, composed of less conserved accessory nucleoporins, are cell-type specific and modifiable by cellular processes.
The main contributors to cytoplasmic FG repeats are Nup214, Nup358, Nup98, and Nup42. Nup358 contributes most of the molecular mass and forms a homopentameric complex that assembles onto the stems of two Y complexes, stabilizing the Y complex rings.
 Cytoplasmic Ring Model
Cytoplasmic Ring Model
4. What Are Nuclear Basket Nucleoporins?
In humans, the nuclear basket is made up of three nucleoporins: Nup50, Nup153, and Tpr. Tpr is the major structural component, containing a large coiled-coil domain that allows for homo-oligomerization.
Nup153 tethers Nup50 to the nuclear pore and recruits Tpr to the NPC post-mitotically. The nuclear basket is critical for mRNA production and quality control, chromosome organization, and DNA damage repair.
5. What Are Karyopherins?
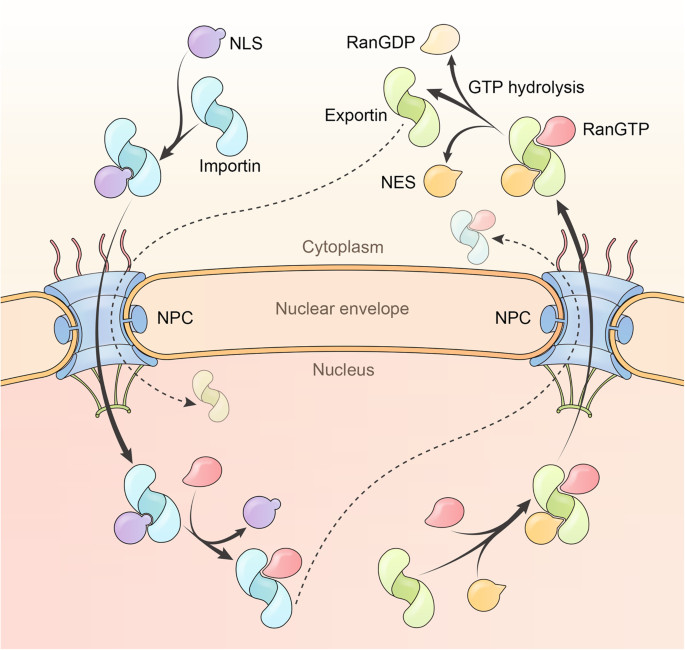
Karyopherins are transport receptors that ferry cargoes across NPCs into or out of the nucleus. They are crucial for various cellular processes and are divided into three groups:
- Importins: Import cargoes into the nucleus.
- Exportins: Export cargoes to the cytoplasm.
- Biportins: Transport cargoes in either direction.
These karyopherins rely on the GTPase RanGTP for cargo binding and dissociation. They are large in size (~ 1000 residues), forming alpha-helical solenoid structures.
5.1. What Are Importins and How Do They Work?
Humans possess ten verified importins, including Importin β1 (Impβ1), Transportin 1 (TNPO1), Transportin 2 (TNPO2), and Transportin 3 (TNPO3). Importins bind to cargoes in the cytoplasm and release them within the nucleus upon encountering RanGTP.
5.1.1. Importin β1
Importin β1 acts as a transporter in classical nuclear import, recruiting the adaptor protein Importin α (Impα), which directly binds to the cargo protein. Impβ1 also imports m3G-caped U snRNA by binding to the IBB of the adaptor snurportin. As a major import receptor, Impβ1 imports many cargoes, including the NF-κB subunit p65, autophagy transcription factor TFEB, and programmed cell death ligand 1 PD-L1.
5.1.2. Transportin 1 and Transportin 2
Transportin 1 (TNPO1) and Transportin 2 (TNPO2) bind the PY (proline-tyrosine) nuclear localization signal (NLS) of cargo proteins. They also bind arginine-glycine-glycine (RGG) domains in RNA binding proteins such as hnRNP A1 and FUS.
5.1.3. Transportin 3
Transportin 3 (TNPO3) specifically binds cargoes containing an arginine/serine (RS) domain. TNPO3 cargoes include alternative splicing factor/splicing factor 2 (ASF/SF2), cold-inducible RNA-binding protein (CIRBP), and polyadenylation specificity factor 6 (CPSF6).
5.2. What Are Exportins and How Do They Work?
The human genome encodes five exportins to conduct the export of cellular proteins and RNAs. Unlike importins, exportins display low affinities to either RanGTP or the cargo alone. However, they can cooperatively bind cargoes and RanGTP and form nanomolar affinity complexes in the nucleus.
5.2.1. Exportin 1
Exportin 1 (XPO1) is the best-characterized exportin with approximately 200 validated protein cargoes, including p53, FOXO, Survivin, TFEB, and cyclic GMP-AMP synthase cGAS. XPO1 interacts with leucine-rich nuclear export signals (NES), which are typically made up of four large hydrophobic residues separated by 1–3 linker residues.
5.2.2. Exportin 2
Exportin 2 (XPO2) is a dedicated nuclear export factor for the classical nuclear import adaptor Impα. By wrapping around RanGTP and Impα, XPO2 ensures cargo dissociation from Impα before export.
5.2.3. Exportin 5
Exportin 5 (XPO5) exports pre-miRNA, a step necessary for proper miRNA maturation. XPO5 recognizes the 2-nucleotide 3’ overhang structure and the double-stranded stem of pre-miRNA.
5.3. What Are Bi-Directional Transporters (Biportins)?
Biportins can function as importins to import cargoes or as exportins to export cargoes. There are three verified biportins in humans:
5.3.1. Importin 13
Importin 13 (IPO13) imports glucocorticoid receptor GR, the exon junction complex components Mago-Y14, the E2 SUMO-conjugating enzyme Ubc9, programmed cell death 5 PDCD5, while exporting translation initiation factor 1 A eIF1A.
5.3.2. Exportin 4
Exportin 4 (XPO4) mediates nuclear import of transcription factors Sox2 and SRY, the glycolytic enzyme PKM2, and nuclear export of Smad proteins, the hypusine-containing translation factor eIF5A, and a subset of circRNAs.
5.3.3. Exportin 7 and RanBP17
Exportin 7 (XPO7) was initially identified as an exportin for RhoGAP1 and 14-3-3sigma. XPO7 can also recognize positively charged folded domains and mediate the nuclear import of NFκB/p65. RanBP17 is 67% identical to XPO7.
5.4. What Are Smaller Size Transport Factors?
These factors do not form α-solenoids but function as karyopherins and can recognize cargoes and translocate through NPCs. Unlike typical karyopherins, they are very specific in cargo recognition, do not rely on RanGTP for cargo binding and dissociation, and contain fewer FG pockets.
5.4.1. Nuclear Transport Factor 2
Nuclear Transport Factor 2 (NTF2) is a dedicated RanGDP nuclear import factor that recycles inactive RanGDP to the nucleus. NTF2 forms a homodimer and uses a distinct hydrophobic cavity for recognition of RanGDP.
5.4.2. NXF1 Family
The nuclear RNA export factor 1 (NXF1) family of proteins possesses an NTF2 domain capable of FG repeat binding and interacts with NTF2-like export factor 1 (NXT1) to form a heterodimer. NXF1 facilitates mRNA nuclear export since it also contains other domains that interact with RNA and other mRNA processing factors.
5.4.3. Hikeshi
The heat shock nuclear import factor Hikeshi mediates nuclear import of molecular chaperone Hsp70 to counteract heat-shock damage and increase cell viability under heat shock conditions.
5.5. What Are Transport Adaptor Molecules?
The transport adaptor itself does not have NPC translocation capabilities; however, it can bind karyopherin and cargo at the same time, thereby facilitating cargo transportation.
5.5.1. Importin α Family
The importin α (Impα) family of adaptors functions in classical nuclear import and recognizes classical NLS signals. Impα contains an N-terminal Importin beta binding (IBB) domain that directly binds to Impβ1.
5.5.2. Snurportin
Snurportin is the nuclear import adaptor for m3G-capped U snRNPs, which participate in pre-mRNA splicing. Snurportin uses an IBB domain to interact with Impβ1.
6. What Are Ran System Proteins?
The transport directionality of importins, exportins, and biportins relies on an elaborate RanGTP system. This system generates the RanGTP gradient, strictly partitioning RanGTP in the nucleus and RanGDP in the cytoplasm.
6.1. How Does Ran Work?
The Ras-related nuclear protein Ran contains a typical small GTPase domain. Ran is active when it is GTP-bound and inactive when it is GDP-bound. RanGTP typically binds importins with nanomolar to picomolar affinities, and its binding induces cargo dissociation.
6.2. How Does RCC1 Work?
Regulator of chromosome condensation (RCC1) directly binds nucleosomal DNA and histones. It collapses the P-loop of Ran to release bound nucleotides and increases guanine nucleotide dissociation.
6.3. How Does RanGAP1 Work?
RanGAP1 is a cytoplasm-localized Ran-specific GAP recruited to the cytoplasmic filament protein Nup358 when SUMOylated. RanGAP1 positions Ran’s catalytic glutamine in the active conformation to trigger hydrolysis.
6.4. How Do RanBP1, RanBP2, RanBP3, and Nup50 Work?
RanBP1 is a coactivator of RanGTP hydrolysis that increases the rate of RanGAP1-mediated RanGTP hydrolysis. RanBP2 (Nup358) contains four RBDs functioning similarly to RanBP1. RanBP3 is a nuclear-localized RBD-containing protein that promotes nuclear export cargo assembly. Basket-localized Nup50 contains a high-affinity importin-binding FG domain and a C-terminal RBD that can recruit RanGTP to accelerate cargo dissociation from importins.
7. How Does Translocation Across the NPC Barrier Work?
Nucleoporins, karyopherins, and Ran system proteins work together to transport cargo through the NPC. Each NPC can transport cargo at a staggering rate of ~ 1000 molecules per second, simultaneously preventing non-specific passive diffusion.
One model proposes that FG nucleoporins bind karyopherins on one side of the NPC, escort them across the barrier, and release them on the other side. However, data suggests that the interaction between FG repeats and karyopherins is rather dynamic, with karyopherins rapidly “sliding” on the FG repeats of different nucleoporins to move forward.
7.1. What Is The Affinity Gradient Model?
This model demands that the trafficking inside the barrier is constrained to a single direction, with the asymmetrically distributed FG types establishing an affinity gradient for karyopherins and luring them toward the high affinity end.
RanGTP and cargo binding regulate the FG pocket size of karyopherin, shaping it selective for certain size balls. After cargo binding, the importin FG pocket enlarges to bind 2 F balls and move along the 2 F gradient to the basket side. In the nucleus, RanGTP binding reshapes the importin FG pocket to select for 1 F balls and drive the importin to the cytoplasmic side.
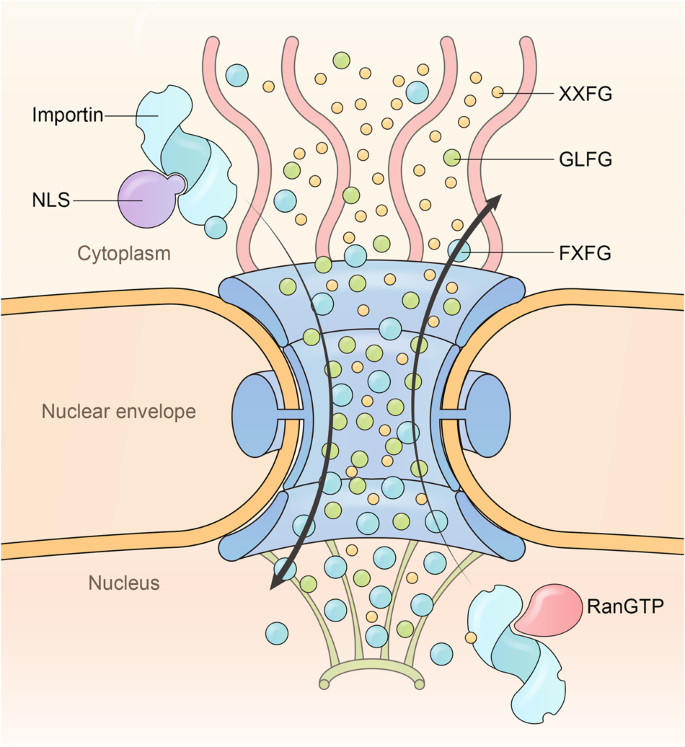 Uni-Directional NPC Translocation Model
Uni-Directional NPC Translocation Model
8. FAQ About Structures That Transport From The Nucleus
8.1. What is the primary function of the Nuclear Pore Complex (NPC)?
The NPC’s primary function is to regulate and facilitate the transport of molecules between the nucleus and the cytoplasm.
8.2. What are the main components of the NPC structure?
The main components include the central core, cytoplasmic filaments, and the nuclear basket.
8.3. What role do nucleoporins play in the NPC?
Nucleoporins form the building blocks of the NPC, contributing to its structural integrity and functional diversity, including anchoring, scaffolding, and forming diffusion barriers.
8.4. How do karyopherins assist in nuclear transport?
Karyopherins act as transport receptors, recognizing and ferrying specific cargoes across the NPC, either into or out of the nucleus.
8.5. What is the significance of FG repeats in nucleoporins?
FG repeats are essential for forming a selective barrier within the NPC, allowing passage of karyopherin-cargo complexes while restricting the entry of other macromolecules.
8.6. What is the function of importins and exportins?
Importins facilitate the transport of molecules into the nucleus, while exportins are responsible for transporting molecules from the nucleus to the cytoplasm.
8.7. How does the Ran system regulate nuclear transport?
The Ran system, with its GTPase activity, establishes a concentration gradient that drives the directionality of nuclear transport, ensuring cargoes are delivered to the correct location.
8.8. What is the affinity gradient model in NPC translocation?
The affinity gradient model suggests that asymmetrically distributed FG types establish an affinity gradient for karyopherins, guiding them toward the high-affinity end, thus facilitating efficient transport.
8.9. How do transport adaptors facilitate nuclear transport?
Transport adaptors bind to karyopherins and cargo simultaneously, facilitating cargo transportation by linking the cargo to the transport machinery.
8.10. Why is the NPC considered a selective transport structure?
The NPC is considered selective because it allows only specific molecules, like karyopherin-cargo complexes, to pass through, while preventing non-specific passive diffusion of other macromolecules.
9. Unlock Insights and Solutions for the Transportation Industry
Want to delve deeper into the fascinating world of transportation structures? Visit worldtransport.net today to explore our extensive collection of articles and resources. Whether you’re seeking detailed analyses, trend forecasts, or innovative solutions, worldtransport.net provides the expertise and insights you need. Contact us at +1 (312) 742-2000 or visit our address at 200 E Randolph St, Chicago, IL 60601, United States, and discover how we can support your journey.
