
Auxin transport involves multiple mechanisms and proteins, creating a robust system that responds to developmental and environmental cues; discover more about this intricate process at worldtransport.net. This ensures plants can adapt to changing conditions and maintain optimal growth patterns. Auxin transporters, including PIN proteins, ABC transporters, and AUX/LAX proteins, work synergistically to regulate auxin flow, influencing various developmental processes. Explore comprehensive insights into plant hormones and efficient nutrient transport systems at worldtransport.net.
1. What Is Auxin And Its Role In Plant Development?
Auxin plays a central role in plant development by mediating the distribution of this critical growth regulator, which influences various processes. Auxin, a unique plant growth substance, controls a wide range of developmental processes through its differential distribution within plants. These non-uniform auxin levels respond to both external stimuli and internal developmental programs, acting as an interface between environmental signals and specific developmental events. To delve deeper into how auxin influences plant morphology, visit worldtransport.net.
The level of any compound in a cell is modulated by two primary processes: metabolism and transport. Both auxin metabolism and transport are crucial in modulating plant development. Multiple transport proteins are essential for maintaining directional auxin flows within plant organs and tissues.
2. What Are The Physical-Chemical Principles Behind Auxin Transport?
The movement of auxin relies on its physical-chemical properties, requiring active transport mechanisms. The distribution of indole-3-acetic acid (IAA), a native auxin, occurs over short and long distances within a plant. While long-distance transport occurs via the vascular system through mass flow, cell-to-cell transport is mostly polar, relying on the physical-chemical nature of auxin molecules. For more information on the vascular system’s role, check out related articles on worldtransport.net.
Auxins are weak acids; their molecular form (proton-dissociated or non-dissociated) determines their ability to penetrate membranes, influenced by pH levels. In plants, the apoplast has a pH of approximately 5.5 due to proton extrusion by plasma membrane H+-ATPases. At this pH, IAA molecules are approximately 83% dissociated (anionic, A−) and 17% proton-associated (HA, i.e., non-dissociated).
Only proton-associated IAA molecules can enter the cell via lipophilic diffusion across the plasma membrane without a carrier protein. In the cytoplasm, with a pH of approximately 7, auxin molecules shift almost entirely to the anionic form. Anionic auxins cannot diffuse across the plasma membrane; thus, cells function as weak acid anion traps unless efflux transporters are present. Asymmetrical localization of these transporters provides directional cellular auxin efflux, establishing polar flows. This concept forms the chemiosmotic polar diffusion model.
3. What Are The Major Types Of Auxin Transporters?
Auxin transporters include PIN proteins, ABC transporters, and AUX/LAX proteins, each playing a unique role in auxin movement. Active transport of auxin anions out of cells requires auxin efflux carriers. There are at least two protein families with auxin-exporting activity: the plant-specific PIN family and the ATP-binding cassette (ABC) superfamily, particularly its B type (ABCB/multidrug resistance/phosphoglycoprotein). These transporters ensure the directional flow of auxin.
3.1. PIN Proteins
PIN proteins were associated with polar auxin transport in the late 1990s. PINs are integral membrane proteins with a topology similar to some transporter proteins. In Arabidopsis, the PIN family consists of eight members divided into two subclades based on the length of their hydrophilic loop. Canonical “long” PINs (PIN1–4 and 7) show mostly polar plasma membrane localization, with strong support for PIN1, 2, 4, and 7 directing auxin transport. Explore the influence of PIN proteins on plant physiology and crop productivity at worldtransport.net.
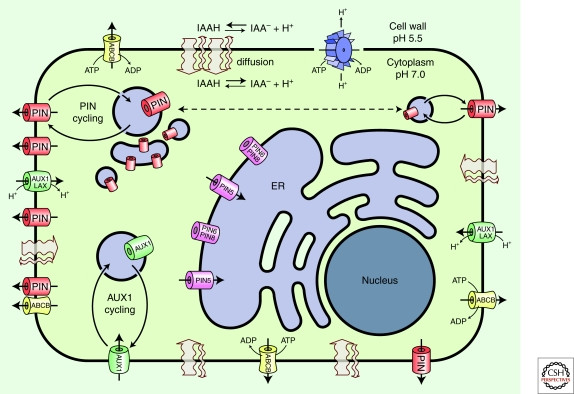 PIN efflux carrier proteins depicted in red represent “long” PINs (PIN1, 2, 3, 4, and 7), whereas PINs marked in pink represent “short” PINs (PIN5, 6, and 8)
PIN efflux carrier proteins depicted in red represent “long” PINs (PIN1, 2, 3, 4, and 7), whereas PINs marked in pink represent “short” PINs (PIN5, 6, and 8)
The polar localization of long PINs determines the direction of auxin flow, influencing many auxin-dependent processes in plant development. These PINs undergo constitutive cycling between the plasma membrane and endosomal compartments, rapidly relocalizing via a transcytosis-like mechanism in response to environmental signals. This polar localization underlies vectorial auxin transport required for embryo development, organogenesis, and tropisms.
In contrast, short PINs (PIN5, 6, and 8) have a reduced hydrophilic loop. PIN5 localizes to the endoplasmic reticulum (ER) and functions as an auxin transporter, suggesting a role in intracellular auxin distribution and regulation of cellular auxin homeostasis. None of the PIN sequences contain an ATP-binding domain, and they are considered secondary transporters, though the energy source for their operation is unknown.
While all PINs directly transport auxins, long PINs exhibit functional redundancy. Unlike ABCB transporters, PIN-like proteins from other kingdoms show little sequence or functional similarity to plant PINs. Plant PINs exhibit about 15% sequence similarity with related proteins from yeasts and even less similarity to bacterial PIN-like proteins.
3.2. ABC Transporters
The ABC superfamily is one of the largest and most ubiquitous transporter families, associated with movements of various small molecules, nutrients, and xenobiotics. Structural analyses indicate high structural conservation between bacterial and eukaryotic ABC transporters. Despite this, ABC transporters exhibit narrow or promiscuous substrate specificity. Learn about the latest advancements in ABC transporter technology via worldtransport.net.
Phylogenetic analyses suggest a small subgroup of the ABCB subclass, represented by ABCB1 and 19 in Arabidopsis, function in auxin transport across plant species. ABCB1/19 are the closest Arabidopsis orthologs of mammalian ABCB1 multidrug resistance transporters but transport auxin with high substrate specificity.
ABCB1/PGP1 and ABCB19/PGP19 act as auxin efflux carriers, while ABCB4 exhibits more complex transport characteristics. ABCB4 functions as an auxin influx transporter under low auxin concentrations and reverses to efflux as auxin concentrations increase, consistent with its role in auxin-dependent root hair elongation. ABCB1, 4, and 19 exhibit stable, primarily nonpolar plasma membrane localization, though more polar localizations may contribute to tissue-specific functions. ABCB19/PGP19 and PIN1 can co-act, with ABCB stabilizing PIN in plasma membrane microdomains and enhancing overall transport specificity.
3.3. AUX/LAX Proteins
Auxin molecules can enter cells passively but are also actively transported via the H+-symport activity of the AUX1/LAX family of plasma membrane permeases. The Arabidopsis genome encodes one AUX1 and three Like AUX1 (LAX1, LAX2, and LAX3) proteins, sharing about 80% amino acid sequence similarity. To understand the complete mechanism of how these proteins act, visit worldtransport.net.
The need for this additional active auxin-uptake process arises in cells requiring high and rapid auxin influx, such as in the lateral root cap where AUX1 plays a major role in redirecting polar auxin streams. While some transporters may be multifunctional and transport substrates other than auxins, the number of transporters involved is striking when considering the central role of auxin transport in programmed and plastic plant development.
4. Why Do Plants Need Overlapping Auxin Transporter Systems?
Overlapping auxin transporter systems provide plants with robustness, flexibility, and adaptability during their development. Three classes of auxin transporters, comprising at least 15 different carriers in A. thaliana, may seem like unnecessary redundancy to a design engineer. However, in biology, a combination of overlapping activities and functional redundancy in a robust transport network is common. Multiple systems often mediate the movement of signaling molecules and vital mineral ions like calcium and nitrate, functioning in specific tissues, cells, and subcellular compartments. For cutting-edge insights into robust transport networks, check out worldtransport.net.
Auxin is mobilized by ATP-driven transporters (ABCBs), H+-symporters (AUX/LAXes), and gradient-driven carriers (PINs). The root of this diversity appears to lie in the improvisational and adaptive processes integral to sessile vascular plant evolution. Mechanisms that control cellular division and expansion were augmented by new means of controlling polar development and the elaboration of complex reproductive/vegetative organs.
Selective pressures favored an integrated network that actuates local responses and transduces a core integrative signal over distance. Vectorial auxin movement provides developmental information, regulated by a localized system of diversion, export, import, and metabolism. Information content is inherent in the signal, position, quantity of flow, and vector of movement rather than a central nervous system. Basal auxin concentrations, localized auxin sources and sinks, and auxin fluxes all direct the position and polarity of growth.
5. What Is The Evolutionary History Of Auxin Transporters?
ABCB transporters are evolutionarily ancient, whereas PIN proteins evolved later with vascular plants, demonstrating the adaptive nature of plants. Phylogenetic analyses of PIN and ABCB transporters indicate that ABCBs constitute a more ancient transport system. ABCB structure is highly conserved throughout phyla, whereas PIN proteins appear to have arisen with vascular plants. Understand how plants adapt and evolve via worldtransport.net.
Endogenous plant flavonoids that emerged in early land plants interfere with the fundamental mechanism of ABCB transporters, whereas PIN proteins are only indirectly affected by these compounds. This suggests that the presence of plant flavonoids was a selective factor in the evolution of PIN1/2-type plasma membrane efflux transporters and provides an evolutionary rationale for overlapping transporter function. Auxin transport must continue to function at basal levels in the presence of flavonoid production in response to herbivory and oxidative stress.
The greater driving force for the elaboration of the PIN efflux system was the more sophisticated developmental programming in vascular plants. The generally apolar localization of ABCB transporters and their high degree of stability on the plasma membrane are not compatible with the plasticity and complexity of development required in terrestrial plants. This includes the evolution of apical growth, vascular tissue, complex organogenesis, and refined polar auxin transport.
6. How Does Functional Analysis Inform Our Understanding Of Auxin Transporters?
Functional analysis reveals how different auxin transporters are used under various conditions, such as in response to energy availability and polarity requirements. ABCB transport is a direct ATP-dependent activity that functions when chemiosmotic gradients are decreased or when auxin must move against a gradient. For example, cortical/epidermal tissues of the mature root maintain auxin flow against the stelar vector to sustain root hair production and other growth phenomena. Discover the newest functional analysis tools via worldtransport.net.
However, a system dependent on direct ATP-driven transport is at a disadvantage in rapidly growing tissues where ATP has been depleted. In these tissues, gradient-driven transporters like PINs and AUX/LAX proteins would be expected to predominate. The need for polarity and plasticity of auxin transport is another important factor. ABCB transporters are highly stable on the plasma membrane and require activation/folding assisted by FKBP42 and other chaperone proteins. Any polarity seen in ABCB localization appears to be associated with secondary cell wall rather than cell plate formation and early events in cytokinesis, and is therefore not dynamic.
In contrast, essential directional polar auxin transport required for embryogenesis and organogenesis is controlled by a subset of PIN proteins trafficked to the plasma membrane with distinct polarized localizations. Plastic growth responses require dynamic redirection of auxin fluxes, with PIN proteins serving as primary vehicles for redirection.
7. How Do Auxin Transporters Coordinate Their Functions?
Auxin transporters coordinate their functions to ensure robust auxin flow, demonstrating the complexity and adaptability of plant transport systems. Simple definitions of auxin transporter function assigned by protein category can be misleading. Overlapping auxin transport mechanisms developed in the context of plant evolution and natural selection result in numerous exceptions. Not all PIN proteins exhibit polar localizations in all tissues, and ABCB localizations are not strictly apolar. Explore adaptive systems in plant transport on worldtransport.net.
Complex interactions of some PIN and ABCB proteins based on indirect interactions further limit this simplistic assignment of function. A better approach defines basic auxin transport streams and views the role of the various transporters in that context.
Developmentally essential polar auxin streams that function in the establishment of embryonic apical–basal polarity and organogenesis appear to be mediated almost exclusively by PIN1, PIN4, and PIN7. Loss of ABCB or AUX1 function has no obvious effect on embryonic development. The auxin flow for lateral root development may be initiated by AUX1 expression in lateral root primordia or relocalization of PIN1 in single protophloem cells. The coordinated action of ABCB1, 19, PIN1, and AUX1 is required for normal lateral root initiation, with AUX1 generating a sink and ABCB1, 19, and PIN1 providing the auxin source.
8. How Do Auxin Transporters Affect Long-Distance Transport Streams?
The loading and restriction of long-distance auxin transport streams rely on the coordinated action of multiple auxin transporters. The long-distance auxin stream from the shoot apical meristem to the root is loaded mainly by PIN1, 7, and ABCB1 and 19. The expression of PIN7 and ABCB1 is more restricted in the shoot apex, consistent with their roles in auxin loading to the vascular stream. The expression of PIN1 and ABCB19 throughout the whole plant maintains the main auxin stream from shoot to root apices. Discover the intricacies of long-distance streams by visiting worldtransport.net.
Acropetal auxin flow (from the mature root to the root tip), driven mainly by PIN1 and ABCB19, is redirected to the lateral root cap and epidermal cells (basipetal flows) through the concerted action of AUX1, ABCB1, 4, 19, and PIN1, 2, 3, and 4. AUX1, ABCB1, 19, and PIN 3,4 function mainly in loading, whereas AUX1, ABCB1, 4, and PIN2 continue the stream along lateral root cap, the epidermal and cortical cells to guide cell division, cell elongation, and root hair development in the outer cell layers.
Rapid auxin influx via AUX1 is consistent with the higher auxin concentration in lateral root cap cells compared with columella cells. Even if ABCB19 and PIN1 appear to be the principal mediators of polar auxin transport along the axis, retention of auxin in vascular transport streams in the shoot is more attributable to the combined export activities of PIN3 and ABCB19, which are localized in bundle sheath cells.
9. How Do Auxin Sinks Drive Transport Streams?
Auxin sinks, created by AUX/LAX proteins, play a crucial role in motivating auxin transport streams, highlighting the importance of these proteins in directional auxin movement. Although auxin transport in the vascular cylinder is primarily associated with xylem parenchyma cells in embryos and seedlings, phloem transport plays a more substantial role in the mature shoot and root. Explore pholem transport and its roles by visiting worldtransport.net.
Recent evidence suggests the importance of auxin sinks generated by AUX/LAX proteins in the motivation of auxin transport streams in the stele cells to generate the auxin fluxes for lateral root development. Mathematical models that have been experimentally verified indicate that AUX1 uptake activity can contribute 10-times more to directional auxin movement than pH-driven passive movement alone.
The contribution of AUX1 in generating auxin sinks is shown by the total loss of gravitropic responses in the aux1 mutant. Loss of function of all other auxin transporters analyzed to date (except PIN2) does not lead to such complete loss of gravitropic growth responses, even when multiple loss-of-function mutations are present. LAX3 plays a similar role in the accumulation of auxin in cortical and epidermal cells surrounding lateral root primordia to facilitate lateral root emergence.
10. How Is Auxin Redirected In Tropic Responses?
Tropic growth responses, such as gravitropism and phototropism, are guided by the dynamic redirection of auxin flow. In response to gravity stimulation, symmetrically localized PIN3 in the root columella relocalizes laterally and facilitates auxin transport to the lateral root cap. The AUX1/PIN2 systems mobilize auxin away from the root apex. In response to unidirectional light, plants redistribute auxin laterally to the distal side with respect to the light, promoting growth there. Learn more about tropic responses by checking out worldtransport.net.
In the Arabidopsis hypocotyl, auxin flow is restricted in the vascular bundle by PIN3 and ABCB19. Unidirectional light triggers its lateral auxin flow, driven by PIN1, 3, and ABCB19. Unlike in the gravitropic response, the relocalization of PIN3 in the hypocotyls of dark-grown seedlings was not seen with unidirectional blue light. Instead, PIN1 at the distal side immediately below the region of phototropic curvature is delocalized, resulting in auxin accumulation above.
11. How Do Competitive Response Mechanisms Affect Auxin Transport?
Competitive response mechanisms, such as those involving nitrate transporters, can influence auxin transport, demonstrating the interconnectedness of nutrient and hormone transport systems. While PINs, ABCBs, and AUX1/LAXes are the primary mediators of auxin transport in plants, other plant anion transporters may exhibit lower levels of auxin transport activity. The apparent induction of auxin uptake and lateral root formation in Arabidopsis grown under low N can be explained by competitive regulation of native auxin transport by nitrate transporters. For a comprehensive overview, visit worldtransport.net.
Similar competitive mechanisms may exist with the malate transporter ABCB14. In all such cases, a localized increase in auxin transport in response to a shortage of the competing small molecule can be readily rationalized.
12. How Do Auxin Transporters Maintain Auxin Homeostasis?
Auxin transporters play a vital role in regulating auxin homeostasis at the cellular level, ensuring optimal auxin concentrations for cellular functions. The ABCB4 transporter functions in this capacity in root trichoblasts. In yeast, ABCB4 has an importer activity at low auxin levels and converts to export activity as auxin levels increase. Such a reversible auxin transporter could maintain auxin homeostasis in root hairs and elongating cells, where constant auxin levels are required and the AUX1 importer is not present. Discover how to optimize plant growth via worldtransport.net.
PIN5, and possibly other “short” PINs (6, 8), which localize to endomembranes (predominantly ER), also play a role in auxin homeostasis. These intracellular auxin transporters change the auxin distribution inside cells, exposing auxin molecules to different conjugating/degrading enzymes. Overexpression of PIN5 results in a dramatic change in the profile of auxin metabolites.
13. How Is Mathematical Modeling Used To Understand Auxin Transport?
Mathematical modeling is a valuable tool for creating hypotheses about the involvement of various transporters in auxin transport, offering insights into complex transport mechanisms. Auxin—with its specialized polar transport features—is well-suited for mathematical modeling. The observation that vascular development in plants is dependent on auxin flux that itself enhances transport capacity of cells further differentiating into vascular bundles led to some of the first mathematical models of the auxin canalization hypothesis. Learn about cutting-edge modeling on worldtransport.net.
Models dealing with the presumption that cells can reinforce their capacity for auxin movement across the plasma membrane by regulation of PIN localization evolved during the last decade. The necessity for PIN protein activity in the creation of auxin maxima responsible for proper phyllotaxis as well as the role of PINs in vein formation have been explored in some detail.
Recent models of phyllotaxis presume that PINs polarize toward the cell with the highest auxin concentration, thus pumping auxin against its concentration gradient and creating auxin maxima in the tissue. The models became more precise with detailed characteristics of auxin carrier abundance on the plasma membrane, taking into account protein cycling, and also auxin-induced PIN expression.
Besides auxin efflux carrier activity, the localization and participation of AUX/LAX influx carriers in auxin accumulation were shown in a model of simplified vein tissue. Despite increasing knowledge about ABCB transporters, they have not been included in mathematical models of auxin flow so far. Interestingly, all current models are still focused on auxin transport at the tissue level and the basic transport element (a single cell) is very simplified there. The advantage of mathematical modeling resides in the possibility to test a number of variants for auxin transport mechanisms, offering a tool to combine complex data sets and test their congruence.
14. What Are Some Frequently Asked Questions About Auxin Transport?
14.1. What Happens If Auxin Transport Is Disrupted?
Disruptions in auxin transport can lead to severe developmental defects, including abnormal organ formation, impaired tropisms, and stunted growth, highlighting the critical role of precise auxin flow in plant development.
14.2. Can Environmental Factors Affect Auxin Transport?
Yes, environmental factors such as light, gravity, and nutrient availability can significantly influence auxin transport by altering the localization and activity of auxin transporters, allowing plants to adapt to changing conditions.
14.3. Are There Any Agricultural Applications Related To Auxin Transport?
Understanding auxin transport mechanisms can lead to agricultural applications, such as optimizing plant architecture for better light capture, improving root development for enhanced nutrient uptake, and developing herbicides that target auxin transport pathways.
14.4. How Does Auxin Transport Differ In Various Plant Species?
Auxin transport mechanisms can vary in different plant species due to differences in the number and types of auxin transporters, as well as variations in regulatory pathways, reflecting the diverse adaptations of plants to their environments.
14.5. What Tools Are Used To Study Auxin Transport?
Researchers use a variety of tools to study auxin transport, including genetic mutants, biochemical assays, imaging techniques, and mathematical modeling, each providing unique insights into the complexities of auxin transport.
14.6. How Does Auxin Interact With Other Plant Hormones During Transport?
Auxin interacts with other plant hormones, such as cytokinin and ethylene, to coordinate various developmental processes. These interactions can influence auxin transport and distribution, leading to synergistic or antagonistic effects.
14.7. What Role Does Phosphorylation Play In Auxin Transport?
Phosphorylation plays a critical role in regulating the activity and localization of PIN proteins, affecting the direction and efficiency of auxin transport. Kinases, such as PINOID, phosphorylate PIN proteins, influencing their polar localization.
14.8. Can Synthetic Auxins Affect Natural Auxin Transport?
Yes, synthetic auxins can affect natural auxin transport by competing with endogenous auxins for transporter binding sites or by altering the expression and localization of auxin transporters, leading to various physiological responses.
14.9. How Is Auxin Transport Related To Apical Dominance?
Auxin transport is closely related to apical dominance, where auxin produced in the apical bud is transported down the stem, inhibiting the growth of lateral buds. Disruptions in auxin transport can weaken apical dominance, leading to increased branching.
14.10. What Are The Ethical Considerations For Researching Auxin Transport?
Ethical considerations for researching auxin transport include avoiding unintended ecological consequences from altering plant hormone pathways, ensuring responsible use of genetically modified plants, and promoting transparency in research findings.
Understanding how auxin is transported is critical for grasping plant development and adaptation. The synergistic action of PIN proteins, ABC transporters, and AUX/LAX proteins, along with their evolutionary and functional complexities, showcases the sophistication of plant transport systems.
Ready to delve deeper into the fascinating world of transport? Visit worldtransport.net today and explore our extensive collection of articles, analyses, and solutions. Whether you’re seeking in-depth insights, trend analyses, or innovative transport solutions, worldtransport.net is your ultimate resource. Contact us at Address: 200 E Randolph St, Chicago, IL 60601, United States. Phone: +1 (312) 742-2000, or visit our Website: worldtransport.net to unlock the full potential of our expertise.