
Serotonin, a crucial neurotransmitter, orchestrates a wide array of functions, from mood regulation and sleep cycles to appetite and even gut motility. Its influence is vast, yet its action is meticulously controlled by a single protein: the serotonin transporter, SERT. This molecular gatekeeper terminates serotonin signaling by diligently回收 (reuptaking) serotonin back into nerve cells, using the energy stored in sodium and chloride gradients. SERT’s pivotal role makes it the primary target for widely prescribed medications treating neuropsychiatric conditions like depression and anxiety. Despite decades of intense research, the precise molecular choreography of serotonin transport by SERT, its intricate coupling with ion gradients, and the enigmatic role of a secondary, allosteric site have remained largely obscure.
Now, groundbreaking research employing cryo-electron microscopy (cryo-EM) has peeled back layers of this mystery. Scientists have captured detailed snapshots of human SERT in action, visualizing it bound to serotonin and in its unbound state, in environments with and without sodium. These structures represent all fundamental stages of the transport cycle, providing unprecedented clarity on how SERT operates. The study pinpoints the location of the allosteric site, revealing it as an aromatic pocket nestled within the scaffold domain in the extracellular vestibule, connected to the primary binding site via a narrow tunnel. These findings offer a transformative structural understanding of allosteric modulation, illuminating how SERT efficiently captures serotonin from the synaptic space and facilitates its release inside neurons.
Two Binding Sites for Serotonin Uncovered in Sodium-Rich Conditions
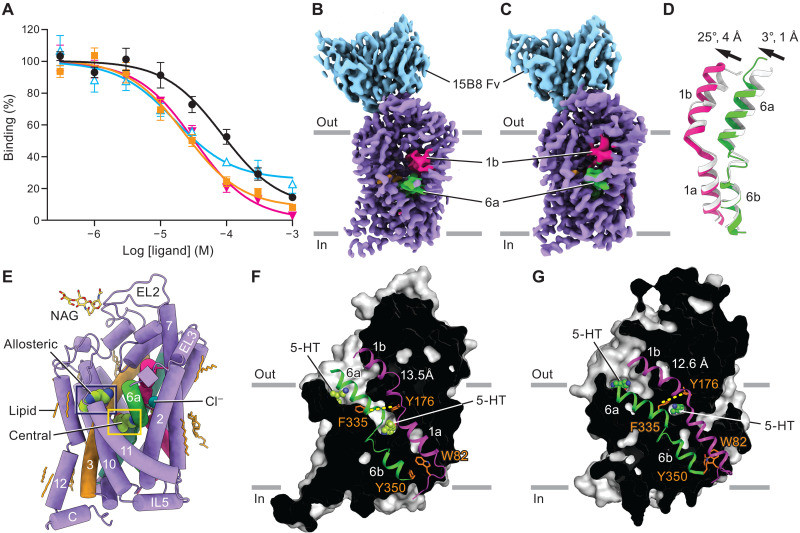
To begin deciphering SERT’s interaction with its cargo, researchers first confirmed that serotonin (5-HT) binds to SERT when it’s embedded in a membrane-like environment (nanodiscs) and in the presence of sodium chloride (NaCl). They used a technique called [3H]paroxetine competition–binding, which measures how well serotonin can displace a known SERT binder (paroxetine). This experiment confirmed serotonin’s binding affinity for SERT under these conditions.
Next, the team employed cryo-EM to visualize the SERT-serotonin complex at near-atomic resolution. Analyzing the 3D images, they identified two distinct conformations of SERT, differing in the extracellular gate region. These conformations were classified as outward-open and occluded, based on the movement of transmembrane helices TM1b and TM6a. This movement dictates whether the central binding site is accessible from the outside of the cell (open) or sealed off (occluded).
Remarkably, in both conformations, transmembrane helix TM1a was found in a closed state, effectively blocking the pathway to the inside of the cell. The outward-open conformation, observed in the majority of particles (87%), closely resembled previously determined structures of SERT bound to antidepressants. The occluded conformation (13% of particles) showed a shift of TM1b and TM6a towards the scaffold domain, resulting in a closed state on both the extracellular and intracellular sides.
Fig. 1. Mechanism of 5-HT capture.
 Fig. 1.
Fig. 1.To further validate their structural findings, the researchers confirmed the locations of sodium (Na+) and chloride (Cl−) ions within the outward-facing conformation. These ions, crucial for driving serotonin transport, were found in positions consistent with previous X-ray structures of SERT. Crucially, the high-resolution cryo-EM maps revealed a clear density for serotonin itself in the central binding site in both outward and occluded conformations.
Detailed analysis showed that serotonin interacts with SERT in a similar manner in both conformations. The tryptamine portion of serotonin nestles between tyrosine residues Tyr176 and Tyr95, while the amine group forms crucial interactions with Ser438 and Asp98. Asp98 is highly conserved among neurotransmitter transporters, highlighting its functional importance. The hydroxyl group of serotonin is positioned between transmembrane helices TM3 and TM8, interacting with Thr439, Ala173, and Ala169. The indole ring of serotonin engages in nonpolar interactions with Ile172 and aromatic interactions with Phe341. Interestingly, Phe335 was observed to undergo a conformational change, moving further into the central binding site as SERT transitions from the outward to the occluded state.
Table 1. Cryo-EM data collection, refinement, and validation statistics.
| 5-HT–SERT outward-NaCl (EMD-23365) (PDB 7LIA ) | 5-HT–SERT occluded-NaCl (EMD-23830) (PDB 7MGW) | 5-HT–SERT inward-KCl (EMD-23364) (PDB 7LI9) | Apo SERT occluded-NaCl (EMD-23362) (PDB 7LI7) | Apo SERT inward-NaCl (EMD-23363) (PDB 7LI8) | Apo SERT inward-KCl (EMD-23361) (PDB 7LI6) |
|—|—|—|—|—|—|—|
| Data collection and processing | 77,160 | 62,383 | 77,160 | 75,642 |
| Magnification | 300 | 300 | 300 | 300 | 300 | 300 |
| Voltage (kV) | 55 | 57 | 50 | 52 | 50 | 53 |
| Electron exposure (e−/Å2) | −0.8 to −2.5 | −0.9 to −2.7 | −1.0 to −2.5 | −1.0 to −2.5 | −1.0 to −2.5 | −1.0 to −2.5 |
| Defocus range (μm) | 0.648 | 0.802 | 0.648 | 0.661 | 0.648 | 0.661 |
| Pixel size (Å) | C1 | C1 | C1 | C1 | C1 | C1 |
| Symmetry imposed | 2,861,741 | 5,624,930 | 5,568,431 | 5,865,511 | 5,551,458 | 5,865,511 |
| Initial particle images (no.) | 774,907 | 114,222 | 299,808 | 636,335 | 343,085 | 707,210 |
| Final particle images (no.) | 3.1 | 3.5 | 3.7 | 3.9 | 3.9 | 3.5 |
| Map resolution (Å) | 0.143 | 0.143 | 0.143 | 0.143 | 0.143 | 0.143 |
| FSC threshold | 3.4–2.6 | 3.8–3.0 | 4.0–3.0 | 4.0–3.2 | 4.2–3.4 | 3.6–3.0 |
| Map resolution range (Å) | | | | | | |
| Refinement | 6DZW | 6DZV | 6DZZ | 6DZV | 6DZZ | 6DZZ |
| Initial model used (PDB code) | 0.59 | 0.78 | 0.83 | 0.56 | 0.74 | 0.64 |
| Initial model CC | 3.5 | 4.0 | 3.9 | 4.1 | 4.2 | 3.8 |
| Model resolution (Å) | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| FSC threshold | −178 | −157 | −161 | −214 | −195 | −185 |
| Map sharpeningB factor (Å2) | | | | | | |
| Model composition | 6,279 | 6,223 | 6,174 | 6,177 | 6,181 | 6,249 |
| Nonhydrogen atoms | 767 | 765 | 767 | 765 | 767 | 767 |
| Protein residues | 231 | 188 | 128 | 142 | 137 | 224 |
| Ligands (atoms) | | | | | | |
| B factors (Å2) | 28 | 192 | 29 | 98 | 99 | 73 |
| Protein | 28 | 180 | 22 | 89 | 91 | 76 |
| Ligand | | | | | | |
| Root mean square deviations | 0.002 | 0.002 | 0.002 | 0.002 | 0.002 | 0.002 |
| Bond lengths (Å) | 0.422 | 0.470 | 0.444 | 0.470 | 0.461 | 0.444 |
| Bond angles (°) | | | | | | |
| Validation | 0.72 | 0.84 | 0.71 | 0.67 | 0.79 | 0.71 |
| Refined model CC | 1.71 | 1.75 | 1.70 | 1.81 | 1.74 | 1.81 |
| MolProbity score | 7.98 | 8.97 | 6.00 | 10.00 | 6.76 | 5.29 |
| Clashscore | 1.71 | 1.40 | 0.00 | 0.00 | 0.00 | 1.72 |
| Poor rotamers (%) | | | | | | |
| Ramachandran plot | 97.50 | 97.10 | 94.61 | 95.78 | 94.74 | 95.01 |
| Favored (%) | 2.50 | 2.90 | 5.39 | 4.22 | 5.26 | 4.99 |
| Allowed (%) | 0 | 0 | 0 | 0 | 0 | 0 |
| Disallowed (%) |
Delving deeper into the central binding site, the researchers observed that serotonin did not completely fill the pocket, suggesting SERT might accommodate slightly modified serotonin molecules. To test this, they examined the binding of serotonin analogs with added methyl groups. Indeed, these analogs showed even higher binding affinity for SERT, supporting the spacious nature of the central site and hinting at SERT’s ability to transport a range of similar molecules.
Intriguingly, careful examination of the cryo-EM maps unveiled an additional density in a hydrophobic pocket within the scaffold domain, present in both outward and occluded conformations. This density, shaped similarly to serotonin in the central site, was identified as a second serotonin molecule bound at an allosteric site. This site is formed by transmembrane helices TM10 to TM12 and extracellular loop 6 (EL6). Key interactions included the hydroxyl group of serotonin projecting between TM10 and TM12, the ligand being cradled by Pro499, and aromatic interactions with Tyr495 and Tyr579. Glu494 forms a hydrogen bond with the amine group of serotonin. Interestingly, Phe556, which occupies this site in unbound SERT, is displaced by serotonin and instead interacts with the amine group from underneath.
Fig. 2. 5-HT binds to central and allosteric sites.
 Fig. 2.
Fig. 2.To validate serotonin binding at this newly identified allosteric site, competition-binding experiments were performed using [3H]vilazodone ([3H]VLZ), an antidepressant known to target the allosteric site. In the presence of imipramine (which occupies the central site), serotonin competed with [3H]VLZ binding, indicating its affinity for the allosteric site. Furthermore, a serotonin analog with a methyl group added to the indole nitrogen, designed to enhance binding at this hydrophobic site, showed a higher binding affinity than serotonin itself, further confirming the location and nature of the allosteric site.
These findings demonstrate that in the presence of sodium, serotonin binds to two distinct locations on SERT: the primary, central binding site, and a secondary, allosteric site. However, unlike antidepressant drugs that bind to the allosteric site and directly block ligand exit from the central site, serotonin binds in a way that does not directly obstruct the central site. The researchers propose that serotonin binding to the allosteric site might indirectly influence ligand unbinding from the central site, possibly by stabilizing the occluded conformation of SERT.
Potassium Induces Inward-Facing State and Serotonin Release
Potassium ions (K+) are known to be counter-transported by SERT and play a role in conformational changes in related transporters. To investigate how serotonin is released into the cell and the allosteric site’s role in this process, the researchers conducted structural and binding studies in the presence of potassium chloride (KCl) and absence of sodium.
Using [3H]ibogaine competition–binding experiments, they found that serotonin binding to SERT was independent of sodium, consistent with previous research. They then determined the cryo-EM structure of the SERT-serotonin complex in KCl. This revealed a single conformation: inward-facing SERT.
Compared to the occluded conformation observed in sodium-rich conditions, the extracellular pathway in the potassium condition was closed even further. Crucially, transmembrane helix TM1a moved “outward,” opening the intracellular gate and removing a barrier (Tyr95) to serotonin release into the cytoplasm.
Fig. 3. Molecular insights into 5-HT release.
 Fig. 3.
Fig. 3.Remarkably, even in this inward-facing conformation, a strong density for serotonin remained in both the central and allosteric sites. This suggests that the allosteric site’s structural integrity is maintained throughout the conformational changes and that serotonin remains bound even after the extracellular gate closes. The presence of serotonin in the allosteric site didn’t seem to hinder extracellular gate closure or the overall conformational transition. The researchers suggest that the allosteric site is compact enough to remain stable even as SERT undergoes these changes.
Further analysis revealed a tunnel connecting the allosteric site to the central site in the outward-facing conformation. This suggests a potential pathway for serotonin to access the central site not only from the extracellular environment but also from the allosteric site. Since the allosteric site remains occupied across different conformations, it could act as a reservoir of serotonin, ready to quickly refill the central site when SERT returns to the outward-facing state, thus accelerating the transport cycle.
Apo-SERT Favors Inward-Facing State in Potassium, Dynamic Behavior in Sodium
To understand SERT’s state when unbound to serotonin, the researchers determined the structure of apo-SERT (serotonin-free SERT) in both potassium and sodium conditions. Preparing apo-SERT involved removing weakly bound serotonin after purification. This apo-SERT preparation was confirmed to be active and ligand-free using binding assays.
Cryo-EM of apo-SERT in KCl revealed a single inward-open conformation. In this state, the extracellular gate is closed, and the central binding pocket is accessible only from the intracellular side. Transmembrane helix TM1a adopted a slightly different, more open conformation compared to the serotonin-bound inward-facing state. This flexibility of TM1a in the inward-open state aligns with molecular dynamics simulations of related transporters, highlighting its dynamic nature.
Fig. 4. Cryo-EM reconstructions of apo SERT in KCl and NaCl.
 Fig. 4.
Fig. 4.These results indicate that in the absence of sodium and presence of potassium, SERT favors an inward-facing conformation with a closed extracellular gate. While the study couldn’t definitively locate potassium ions in the structure, it suggests that either potassium directly stabilizes the inward-open state or, more simply, the absence of sodium drives this conformation.
In contrast, examining apo-SERT in sodium-rich conditions revealed a more complex picture. Cryo-EM identified two distinct conformations: occluded and inward-open. In the occluded state (about 55% of particles), TM1a is closed, preventing access to the cytoplasm. In the inward-open state (about 30%), TM1a is open, allowing intracellular access. Both conformations had a closed extracellular gate. The occluded state of apo-SERT in sodium was virtually identical to the serotonin-bound occluded state.
The presence of multiple conformations of apo-SERT in sodium suggests that, without serotonin bound, SERT exists in a dynamic equilibrium, sampling various states, potentially including outward-open states. The researchers propose that sodium binding to apo-SERT creates this dynamic behavior, priming the transporter for serotonin capture. When SERT transiently adopts an outward-facing conformation and serotonin binds to the central site (possibly aided by the allosteric site), the transport cycle is initiated.
Proposed Mechanism of Serotonin Transport by SERT
The identification of the allosteric site for serotonin, distinct from the previously known allosteric sites for antidepressants, sheds new light on SERT regulation. This study provides the first structural evidence of serotonin occupying an allosteric site in SERT. This site, while close to the antidepressant binding site, does not directly block the central site.
The researchers propose a refined model for serotonin transport by SERT, based on their cryo-EM structures:
-
Outward-Facing State: SERT starts in an outward-facing conformation, bound to sodium and chloride ions. Serotonin can bind to the central site, potentially facilitated by initial capture at the allosteric site.
-
Occluded State: SERT transitions to an occluded conformation, closing off both extracellular and intracellular pathways. The allosteric site, if occupied, retains serotonin.
-
Inward-Facing State: SERT shifts to an inward-facing conformation, opening the intracellular gate and releasing serotonin and a sodium ion into the cytoplasm.
-
Return to Outward-Facing: SERT cycles back to the outward-facing conformation, likely through an equilibrium of states. In the outward-facing state, the central site can re-bind serotonin, potentially drawing from the allosteric site reservoir, to initiate another transport cycle.
Fig. 5. 5-HT transport mechanism of SERT.
 Fig. 5.
Fig. 5.This detailed structural understanding of the Serotonin Transporter Sert mechanism, including the role of the allosteric site and conformational changes, provides a crucial foundation for developing more targeted and effective therapies for neuropsychiatric disorders. By visualizing the molecular dance of serotonin transport, this research opens new avenues for drug design and a deeper understanding of brain function.
References
The references are provided in the original article. Please refer to the “[bài viết gốc]” for the complete list.
